§ V. ANOTHER EXAMPLE OF LIFE WITHOUT AIR—FERMENTATION OF LACTATE OF LIME
AS another example of life without air, accompanied by fermentation properly so called, we may lastly cite the fermentation of lactate of lime in a mineral medium.
In the experiment described in the last paragraph, it will be remembered that the ferment liquid and the germs employed in its impregnation came in contact with air, although only for a very brief time. Now, notwithstanding that we possess exact observations which prove that the diffusion of oxygen and nitrogen in a liquid absolutely deprived of air, so far from taking place rapidly, is, on the contrary, a very slow process indeed; yet we were anxious to guard the experiment that we are about to describe from the slightest possible trace of oxygen at the moment of impregnation.
We employed a liquid prepared as follows: Into from 9 to 10 litres (somewhat over 2 gallons) of pure water the following salts〖Should the solution of lactate of lime be turbid, it may be clarified by filtration, after previously adding a small quantity of phosphate of ammonia, which throws down phosphate of lime. It is only after this process of clarification and filtration that the phosphates of the formula are added. The solution soon becomes turbid if left in contact with air, in consequence of the spontaneous formation of bacteria.〗 were introduced successively, viz:
Pure lactate of lime………………225 grammesPhosphate of ammonia………………0.75 “Phosphate of potassium…………0.4 “Sulphate of magnesium………………0.4 “Sulphate of ammonia……………………0.2 “
(1 gramme = 15.43 grains)
On March 23rd, 1875, we filled a 6 litre (about 11 pints) flask, of the shape represented in and placed it over a heater. Another flame was placed below a vessel containing the same liquid, into which the curved tube of the flask plunged. The liquids in the flask and in the basin were raised to boiling together, and kept in this condition for more than half-an-hour, so as to expel all the air held in solution. The liquid was several times forced out of the flask by the steam, and sucked back again; but the portion which re-entered the flask was always boiling. On the following day when the flask had cooled, we transferred the end of the delivery tube to a vessel full of mercury and placed the whole apparatus in an oven at a temperature varying between 25° C. and 30° C. (77° F. and 86° F.); then, after having refilled the small cylindrical tap-funnel with carbonic acid, we passed into it with all necessary precautions 10 cc. (0.35 fl. oz.) of a liquid similar to that described, which had been already in active fermentation for several days out of contact with air and now swarmed with vibrios. We then turned the tap of the funnel, until only a small quantity of liquid was left, just enough to prevent the access of air. In this way the impregnation was accomplished without either the ferment-liquid or the ferment-germs having been brought in contact, even for the shortest space, with the external air. The fermentation, the occurrence of which at an earlier or later period depends for the most part on the condition of the impregnating germs, and the number introduced in the act, in this case began to manifest itself by the appearance of minute bubbles from March 29th. But not until April 9th did we observe bubbles of larger size rise to the surface. From that date onward they continued to come in increasing number, from certain points at the bottom of the flask, where a deposit of earthy phosphates existed; and at the same time the liquid, which for the first few days remained perfectly clear, began to grow turbid in consequence of the development of vibrios. It was on the same day that we first observed a deposit on the sides of carbonate of lime in crystals.
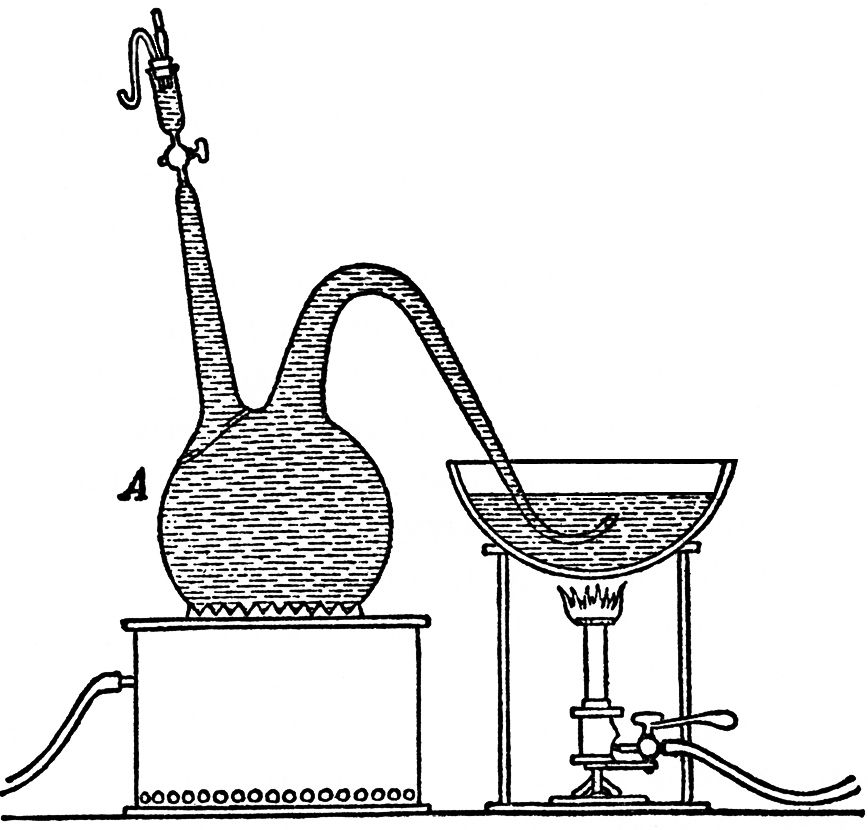
FIG. 11
It is a matter of some interest to notice here that, in the mode of procedure adopted, everything combined to prevent the interference of air. A portion of the liquid expelled at the beginning of the experiment, partly because of the increased temperature in the oven and partly also by the force of the gas, as it began to be evolved from the fermentative action, reached the surface of the mercury, where, being the most suitable medium we know for the growth of bacteria, it speedily swarmed with these organisms.〖The naturalist Cohn, of Breslau, who published an excellent work on bacteria in 1872, described, after Mayer, the composition of a liquid peculiarly adapted to the propagation of these organisms, which it would be well to compare for its utility in studies of this kind with our solution of lactate and phosphates. The following is Cohn's formula:
Distilled water …………………………………………20 cc. (0.7 fl. oz.)Phosphate of potassium ………………………0.1 gramme (1.5 grains)Sulphate of magnesium …………………………0.1 “Tribasic phosphate of lime ……………0.01 (0.15 grain)Tartrate of ammonia ………………………………0.2 (3 grains)
This liquid, the author says, has a feeble acid reaction and forms a perfectly clear solution.〗 In this way any passage of air, if such a thing were possible, between the mercury and the sides of the delivery-tube was altogether prevented, since the bacteria would consume every trace of oxygen which might be dissolved in the liquid lying on the surface of the mercury. Hence it is impossible to imagine that the slightest trace of oxygen could have got into the liquid in the flask.
Before passing on we may remark that in this ready absorption of oxygen by bacteria we have a means of depriving fermentable liquids of every trace of that gas with a facility and success equal or even greater than by the preliminary method of boiling. Such a solution as we have described, if kept at summer heat, without any previous boiling, becomes turbid in the course of twenty-four hours from a spontaneous development of bacteria; and it is easy to prove that they absorb all the oxygen held in solution.〖On the rapid absorption of oxygen by bacteria, see also our Mémoire of 1872, surles Générations dites Spontanées, especially the note on page 78.〗 If we completely fill a flask of a few litres capacity (about a gallon) (FIG. 9 with the liquid described, taking care to have the delivery-tube also filled, and its opening plunged under mercury, and, forty-eight hours afterwards by means of a chloride of calcium bath, expel from the liquid on the surface of the mercury all the gas which it holds in solution, this gas, when analyzed, will be found to be composed of a mixture of nitrogen and carbonic acid gas, without the least trace of oxygen. Here, then, we have an excellent means of depriving the fermentable liquid of air; we simply have completely to fill a flask with the liquid, and place it in the oven, merely avoiding any addition of butyric vibrios, before the lapse of two or three days. We may wait even longer; and then, if the liquid does become impregnated spontaneously with vibrio germs, the liquid, which at first was turbid from the presence of bacteria, will become bright again, since the bacteria, when deprived of life, or, at least, of the power of moving, after they have exhausted all the oxygen in solution, will fall inert to the bottom of the vessel. On several occasions we have determined this interesting fact, which tends to prove that the butyric vibrios cannot be regarded as another form of bacteria, inasmuch as, on the hypothesis of an original relation between the two productions, butyric fermentation ought in every case to follow the growth of bacteria.
We may also call attention to another striking experiment, well suited to show the effect of differences in the composition of the medium upon the propagation of microscopic beings. The fermentation which we last described commenced on March 27th and continued until May 10th; that to which we are now to refer, however, was completed in four days, the liquid employed being similar in composition and quantity to that employed in the former experiment. On April 23, 1875, we filled a flask of the same shape as that represented in and of similar capacity, viz., 6 litres, with a liquid composed as described at page 324. This liquid had been previously left to itself for five days in large open flasks, in consequence of which it had developed an abundant growth of bacteria. On the fifth day a few bubbles, rising from the bottom of the vessels, at long intervals, betokened the commencement of butyric fermentation, a fact, moreover, confirmed by the microscope, in the appearance of the vibrios of this fermentation in specimens of the liquid taken from the bottom of the vessels, the middle of its mass, and even in the layer on the surface that was swarming with bacteria. We transferred the liquid so prepared to the 6-litre flask arranged over the mercury. By evening a tolerably active fermentation had begun to manifest itself. On the 24th this fermentation was proceeding with astonishing rapidity, which continued during the 25th and 26th. During the evening of the 26th it slackened, and on the 27th all signs of fermentation had ceased. This was not, as might be supposed, a sudden stoppage due to some unknown cause; the fermentation was actually completed, for when we examined the fermented liquid on the 28th we could not find the smallest quantity of lactate of lime. If the needs of industry should ever require the production of large quantities of butyric acid, there would, beyond doubt, be found in the preceding fact valuable information in devising an easy method of preparing that product in abundance.〖In what way are we to account for so great a difference between the two fermentations that we have just described? Probably it was owing to some modification effected in the medium by the previous life of the bacteria, or to the special character of the vibrios used in impregnation. Or, again, it might have been due to the action of the air, which, under the conditions of our second experiment, was not absolutely eliminated, since we took no precaution against its introduction at the moment of filling our flask, and this would tend to facilitate the multiplication of anaërobian vibrios, just as, under similar conditions, would have been the case if we had been dealing with a fermentation by ordinary yeast.〗
Before we go any further, let us devote some attention to the vibrios of the preceding fermentations.
On May 27th, 1862, we completely filled a flask capable of holding 2.780 litres (about five pints) with the solution of lactate and phosphates.〖In this case the liquid was composed as follows: A saturated solution of lactate of lime, at a temperature of 25° C. (77° F.), was prepared, containing for every 100 cc. (3 1/2 fl. oz.) 25.65 grammes (394 grains) of the lactate, C6H5O5CaO (new notation, C6H10CaO6). This solution was rendered very clear by the addition of 1 gramme of phosphate of ammonia and subsequent filtration. For a volume of 8 litres (14 pints) of this clear saturated solution we used (1 gramme=15.43 grains):
Phosphate of ammonia ……………………2 grammesPhosphate of potassium ………………1 “Phosphate of magnesium ………………1 “Sulphate of ammonia ………………………0.5 “〗 We refrained from impregnating it with any germs. The liquid became turbid from a development of bacteria and then underwent butyric fermentation. By June 9th the fermentation had become sufficiently active to enable us to collect in the course of twenty-four hours, over mercury, as in all our experiments, about 100 cc. (about 6 cubic inches) of gas. By June 11th, judging from the volume of gas liberated in the course of twenty-four hours, the activity of the fermentation had doubled. We examined a drop of the turbid liquid. Here are the notes accompanying the sketch as they (FIG. 12) stand in our note-book: “A swarm of vibrios, so active in their movements that the eye has great difficulty in following them.
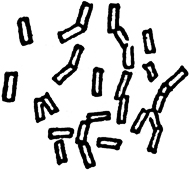
FIG. 12
They may be seen in pairs throughout the field, apparently making efforts to separate from each other. The connection would seem to be by some invisible, gelatinous thread, which yields so far to their efforts that they succeed in breaking away from actual contact, but yet are, for a while, so far restrained that the movements of one have a visible effect on those of the other. By and by, however, we see a complete separation effected, and each moves on its separate way with an activity greater than it ever had before.”
One of the best methods that can be employed for the microscopical examination of these vibrios, quite out of contact with air, is the following. After butyric fermentation has been going on for several days in a flask (FIG. 13), we connect this flask by an india-rubber tube with one of the flattened bulbs previously described, which we then place on the stage of the microscope (FIG. 13). When we wish to make an observation we close, under the mercury, at the point b, the end of the drawn-out and bent delivery-tube. The continued evolution of gas soon exerts such a pressure within the flask, that when we open the tap r, the liquid is driven into the bulb ll, until it becomes quite full and the liquid flows over into the glass V. In this manner we may bring the vibrios under observation without their coming into contact with the least trace of air, and with as much success as if the bulb, which takes the place of an object glass, had been plunged into the very centre of the flask. The movements and fissiparous multiplication of the vibrios may thus be seen in all their beauty, and it is indeed a most interesting sight. The movements do not immediately cease when the temperature is suddenly lowered, even to a considerable extent, 15° C. (59° F.) for example; they are only slackened. Nevertheless, it is better to observe them at the temperatures most favourable to fermentation, even in the oven where the vessels employed in the experiment are kept at a temperature between 25° C. and 30° C. (77° F. and 86° F.).
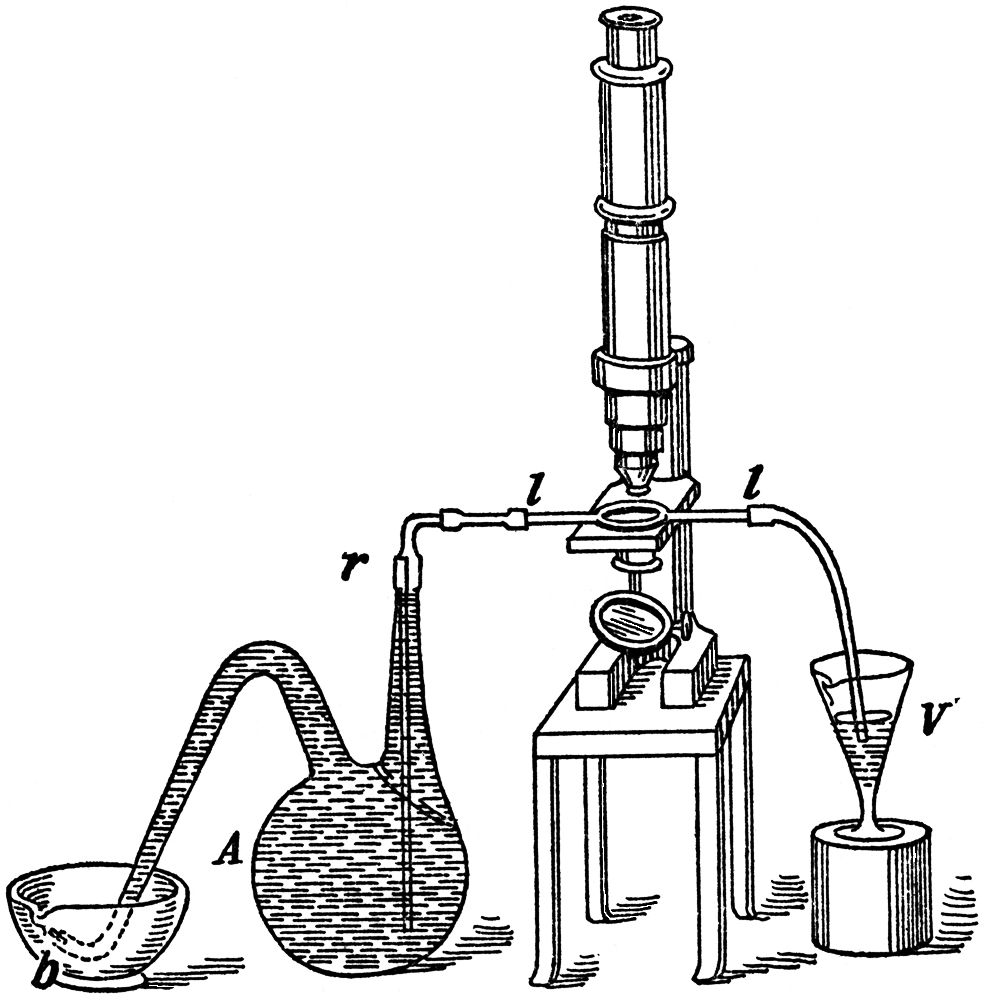
FIG. 13
We may now continue our account of the fermentation which we were studying when we made this last digression. On June 17th that fermentation produced three times as much gas as it did on June 11th, when the residue of hydrogen, after absorption by potash, was 72.6 per cent.; whilst on the 17th it was only 49.2 per cent. Let us again discuss the microscopic aspect of the turbid liquid at this stage. Appended is the sketch we made (FIG. 14) and our notes on it: “A most beautiful object: vibrios all in motion, advancing or undulating. They have grown considerably in bulk and length since the 11th; many of them are joined together in long sinuous chains, very mobile at the articulations, visibly less active and more wavering in proportion to the number that go to form the chain, of the length of the individuals.” This description is applicable to the majority of the vibrios which occur in cylindrical rods and are homogeneous in aspect. There are others, of rare occurrence in chains, which have a clear corpuscle, that is to say, a portion more refractive than other parts of the segments, at one of their extremities. Sometimes the foremost segment has the corpuscle at one end, sometimes the other. The long segments of the commoner kind attain a length of from 10 to 30 and even 45 thousandths of a millimetre. Their diameter is from 1 1/2 to 2, very rarely 3, thousandths of a millimetre.〖1 millimetre=0.039 inch: hence the dimensions indicated will be—length, from 0.00039 to 0.00117, or even 0.00176 in.; diameter, from 0.000058 to 0.000078, rarely 0.000117 in.—D. C. R.〗
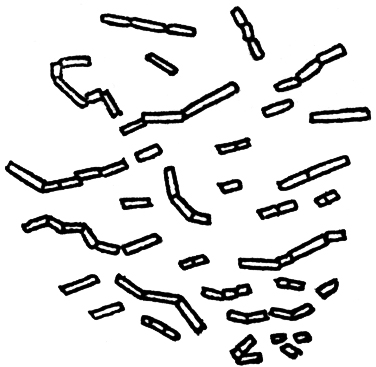
FIG. 14
On June 28th, fermentation was quite finished; there was no longer any trace of gas, nor any lactate in solution. All the infusoria were lying motionless at the bottom of the flask. The liquid clarified by degrees, and in the course of a few days became quite bright. Here we may inquire, were these motionless infusoria, which from complete exhaustion of the lactate, the source of the carbonaceous part of their food, were now lying inert at the bottom of the fermenting vessel—were they dead beyond the power of re vival?(The carbonaceous supply, as we remarked, had failed them, and to this failure the absence of vital action, nutrition, and multiplication was attributable. The liquid, however, contained butyrate of lime, a salt possessing properties similar to those of the lactate. Why could not this salt equally well support the life of the vibrios? The explanation of the difficulty seems to us to lie simply in the fact that lactic acid produces heat by its decomposition, whilst butyric acid does not, and the vibrios seem to require heat during the chemical process of their nutrition.) The following experiment leads us to believe that they were not perfectly lifeless, and that they might behave in the same manner as the yeast of beer, which, after it has decomposed all the sugar in a fermentable liquid, is ready to revive and multiply in a fresh saccharine medium. On April 22nd, 1875, we left in the oven at a temperature of 25° C. (77° F.) a fermentation of lactate of lime that had been completed. The delivery tube of the flask A (FIG. 15), in which it had taken place, had never been withdrawn from under the mercury. We kept the liquid under observation daily, and saw it gradually become brighter; this went on for fifteen days. We then filled a similar flask, B, with the solution of lactate, which we boiled, not only to kill the germs of vibrios which the liquid might contain, but also to expel the air that it held in solution. When the flask, B, had cooled, we connected the two flasks, avoiding the introduction of air, (To do this it is sufficient, first, to fill the curved ends of the stopcocked tubes of the flasks, as well as the india-rubber tube c c which connects them, with boiling water that contains no air.) after having slightly shaken the flask, A, to stir up the deposit at the bottom. There was then a pressure due to carbonic acid at the end of the delivery tube of this latter flask, at the point a, so that on opening the taps r and s, the deposit at the bottom of flask A was driven over into flask B, which in consequence was impregnated with the deposit of a fermentation that had been completed fifteen days before. Two days after impregnation the flask B began to show signs of fermentation. It follows that the deposit of vibrios of a completed butyric fermentation may be kept, at least for a certain time, without losing the power of causing fermentation. It furnishes a butyric ferment, capable of revival and action in a suitable fresh fermentable medium.
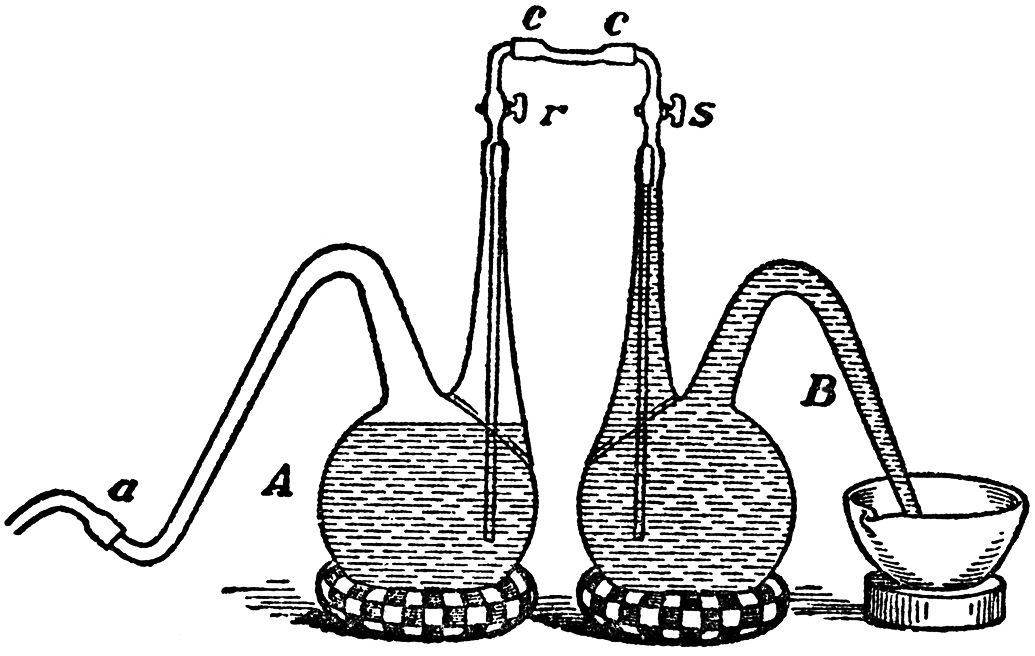
FIG. 15
The reader who has attentively studied the facts which we have placed before him cannot, in our opinion, entertain the least doubt on the subject of the possible multiplication of the vibrios of a fermentation of lactate of lime out of contact with atmospheric oxygen. If fresh proofs of this important proposition were necessary, they might be found in the following observations, from which it may be inferred that atmospheric oxygen is capable of suddenly checking a fermentation produced by butyric vibrios, and rendering them absolutely motionless, so that it cannot be necessary to enable them to live. On May 7th, 1862, we placed in the oven a flask holding 2.580 litres (4 1/2 pints), and filled with the solution of lactate of lime and phosphates, which we had impregnated on the 9th with two drops of a liquid in butyric fermentation. In the course of a few days fermentation declared itself: on the 18th it was active; on the 30th it was very active. On June 1st it yielded hourly 35 cc. (2.3 cubic inches) of gas, containing ten per cent. of hydrogen. On the 2nd we began the study of the action of air on the vibrios of this fermentation. To do this we cut off the delivery-tube on a level with its point of junction to the flask, then with a 50 cc. pipette we took out that quantity (1 3/4 fl. oz.) of liquid which was, of course, replaced at once by air. We then reversed the flask with the opening under the mercury, and shook it every ten minutes for more than an hour. Wishing to make sure, to begin with, that the oxygen had been absorbed we connected under the mercury the beak of the flask by means of a thin india-rubber tube filled with water, with a small flask, the neck of which had been drawn out and was filled with water; we then raised the large flask with the smaller kept above it. A Mohr's clip, which closed the india-rubber tube, and which we then opened, permitted the water contained in the small flask to pass into the large one, whilst the gas, on the contrary, passed upwards from the large flask into the small one. We analyzed the gas immediately, and found that, allowing for the carbonic acid and hydrogen, it did not contain more than 14.2 per cent. of oxygen, which corresponds to an absorption of 6.6 cc., or of 3.3 cc. (0.2 cubic inch) of oxygen for the 50 cc. (3.05 cubic inches) of air employed. Lastly, we again established connection by an india-rubber tube between the flasks, after having seen by microscopical examination that the movements of the vibrios were very languid. Fermentation had become less vigorous without having actually ceased, no doubt because some portions of the liquid had not been brought into contact with the atmospheric oxygen, in spite of the prolonged shaking that the flask had undergone after the introduction of the air. Whatever the cause might have been, the significance of the phenomenon is not doubtful. To assure ourselves further of the effect of air on the vibrios, we half filled two test tubes with the fermenting liquid taken from another fermentation which had also attained its maximum of intensity, into one of which we passed a current of air, into the other carbonic acid gas. In the course of half an hour, all the vibrios in the aerated tube were dead, or at least motionless, and fermentation had ceased. In the other tube, after three hours' exposure to the effects of the carbonic acid gas, the vibrios were still very active, and fermentation was going on.
There is a most simple method of observing the deadly effect of atmospheric air upon vibrios. We have seen in the microscopical examination made by means of the apparatus represented in FIG. 13 how remarkable were the movements of the vibrios when absolutely deprived of air, and how easy it was to discern them. We will repeat this observation, and at the same time make a comparative study of the same liquid under the microscope in the ordinary way, that is to say, by placing a drop of the liquid on an object-glass, and covering it with a thin glass slip, a method which must necessarily bring the drop into contact with air, if only for a moment. It is surprising what a remarkable difference is observed immediately between the movements of the vibrios in the bulb and those under the glass. In the case of the latter, we generally see all movement at once cease near the edges of the glass, where the drop of liquid is in direct contact with the air; the movements continue for a longer or shorter time about the centre, in proportion as the air is more or less intercepted by the vibrios at the circumference of the liquid. It does not require much skill in experiments of this kind to enable one to see plainly that immediately after the glass has been placed on the drop, which has been affected all over by atmospheric air, the whole of the vibrios seem to languish and to manifest symptoms of illness—we can think of no better expression to explain what we see taking place—and that they gradually recover their activity about the centre, in proportion as they find themselves in a part of the medium that is less affected by the presence of oxygen.
Some of the most curious facts are to be found in connection with an observation, the correlative and inverse of the foregoing, on the ordinary aërobian bacteria. If we examine below the microscope a drop of liquid full of these organisms under a coverslip, we very soon observe a cessation of motion in all the bacteria which lie in the central portion of the liquid, where the oxygen rapidly disappears to supply the necessities of the bacteria existing there; whilst, on the other hand, near the edges of the cover-glass the movements are very active, in consequence of the constant supply of air. In spite of the speedy death of the bacteria beneath the centre of the glass, we see life prolonged there if by chance a bubble of air has been enclosed. All round this bubble a vast number of bacteria collect in a thick, moving circle, but as soon as all the oxygen of the bubble has been absorbed they fall apparently lifeless, and are scattered by the movement of the liquid.〖We find this fact, which we published as long ago as 1863, confirmed in a work of H. Hoffmann's, published in 1869 under the title of Mémoire sur les bactéries, which has appeared in French (Annales des Sciences naturelles, 5th series, vol. ix.). On this subject we may cite an observation that has not yet been published. Aërobian bacteria lose all power of movement when suddenly plunged into carbonic acid gas; they recover it, however, as if they had only been suffering from anæsthesia, as soon as they are brought into the air again.〗
We may here be permitted to add, as a purely historical matter, that it was these two observations just described, made successively one day in 1861, on vibrios and bacteria, that first suggested to us the idea of the possibility of life without air, and caused us to think that the vibrios which we met so frequently in our lactic fermentations must be the true butyric ferment.
We may pause to consider an interesting question in reference to the two characters under which vibrios appear in butyric fermentations. What is the reason that some vibrios exhibit refractive corpuscles, generally of a lenticular form, such as we see in We are strongly inclined to believe that these corpuscles have to do with a special mode of reproduction in the vibrios, common alike to the anaërobian forms which we are studying, and the ordinary aërobian forms in which also the corpuscles of which we are speaking may occur. The explanation of the phenomenon, from our point of view, would be that, after a certain number of fissiparous generations, and under the influence of variations in the composition of the medium, which is constantly changing through fermentation as well as through the active life of the vibrios themselves, cysts, which are simply the refractive corpuscles, form along them at different points. From these gemmules we have ultimately produced vibrios, ready to reproduce others by the process of transverse division for a certain time, to be themselves encysted, later on. Various observations incline us to believe that, in their ordinary form of minute, soft, exuberant rods, the vibrios perish when submitted to desiccation, but when they occur in corpuscular or encysted form they possess unusual powers of resistance and may be brought to the state of dry dust and be wafted about by winds. None of the matter which surrounds the corpuscle or cyst seems to take part in the preservation of the germ, when the cyst is formed, for it is all re-absorbed, gradually leaving the cyst bare. The cysts appear as masses of corpuscles, in which the most practiced eye cannot detect anything of an organic nature, or anything to remind one of the vibrios which produced them; nevertheless, these minute bodies are endowed with a latent vital action, and only await favourable conditions to develop long rods of vibrios. We are not, it is true, in a position to adduce any very forcible proofs in support of these opinions. They have been suggested to us by experiments, none of which, however, have been absolutely decisive in their favour. We may cite one of our observations on this subject.
In a fermentation of glycerine in a mineral medium—the glycerine was fermenting under the influence of butyric vibrios—after we had determined the, we may say, exclusive presence of lenticular vibrios, with refractive corpuscles, we observed the fermentation, which for some unknown reason had been very languid, suddenly become extremely active, but now through the influence of the ordinary vibrios. The gemmules with brilliant corpuscles had almost disappeared; we could see but very few, and those now consisted of the refractive bodies alone, the bulk of the vibrios accompanying them having undergone some process of re-absorption.
Another observation which still more closely accords with this hypothesis is given in our work on silk-worm disease (vol. 1, p. 256). We there demonstrated that, when we place in water some of the dust formed of desiccated vibrios, containing a host of these refractive corpuscles, in the course of a very few hours large vibrios appear, well-developed rods fully grown, in which the brilliant points are absent; whilst in the water no process of development from smaller vibrios is to be discerned, a fact which seems to show that the former had issued fully grown from the refractive corpuscles, just as we see colpoda issue with their adult aspect from the dust of their cysts. This observation, we may remark, furnishes one of the best proofs that can be adduced against the spontaneous generation of vibrios or bacteria, since it is probable that the same observation applies to bacteria. It is true that we cannot say of mere points of dust examined under the microscope, that one particular germ belongs to vibrio, another to bacterium; but how is it possible to doubt that the vibrios issue, as we see them, from an ovum of some kind, a cyst, or germ, of determinate character, when, after having placed some of those indeterminate motes of dust into clean water, we suddenly see, after an interval of not more than one or two hours, an adult vibrio crossing the field of the microscope, without our having been able to detect any intermediate state between its birth and adolescence?
It is a question whether differences in the aspect and nature of vibrios, which depend upon their more or less advanced age, or are occasioned by the influence of certain conditions on the medium in which they propagate, do not bring about corresponding changes in the course of the fermentation and the nature of its products. Judging at least from the variations in the proportions of hydrogen and carbonic acid gas produced in butyric fermentations, we are inclined to think that this must be the case; nay, more, we find that hydrogen is not even a constant product in these fermentations. We have met with butyric fermentations of lactate of lime which did not yield the minutest trace of hydrogen, or anything besides carbonic acid. FIG. 16 represents the vibrios which we observed in a fermentation of this kind. They present no special features. Butyl alcohol is, according to our observations, an ordinary product, although it varies and is by no means a necessary concomitant of these fermentations. It might be supposed, since butylic alcohol may be produced, and hydrogen be in deficit, that the proportion of the former of these products would attain its maximum when the latter assumed a minimum. This, however, is by no means the case; even in those few fermentations that we have met with in which hydrogen was absent, there was no formation of butylic alcohol.

FIG. 16
From a consideration of all the facts detailed in this section we can have no hesitation in concluding that, on the one hand, in cases of butyric fermentation, the vibrios which abound in them and constitute their ferment, live without air or free oxygen; and that, on the other hand, the presence of gaseous oxygen operates prejudicially against the movements and activity of those vibrios. But now, does it follow that the presence of minute quantities of air brought into contact with a liquid undergoing butyric fermentation would prevent the continuance of that fermentation or even exercise any check upon it? We have not made any direct experiments upon this subject; but we should not be surprised to find that, so far from hindering, air may, under such circumstances, facilitate the propagation of the vibrios and accelerate fermentation. This is exactly what happens in the case of yeast. But how could we reconcile this, supposing it were proved to be the case, with the fact just insisted on as to the danger of bringing the butyric vibrios into contact with air? It may be possible that life without air results from habit, whilst death through air may be brought about by a sudden change in the conditions of the existence of the vibrios. The following remarkable experiment is well-known: A bird is placed in a glass jar of one or two litres (60 to 120 cubic inches) in capacity which is then closed. After a time the creature shows every sign of intense uneasiness and asphyxia long before it dies; a similar bird of the same size is introduced into the jar; the death of the latter takes place instantaneously, whilst the life of the former may still be prolonged under these conditions for a considerable time, and there is no difficulty even in restoring the bird to perfect health by taking it out of the jar. It seems impossible to deny that we have here a case of the adaptation of an organism to the gradual contamination of the medium; and so it may likewise happen that the anaërobian vibrios of a butyric fermentation, which develop and multiply absolutely without free oxygen, perish immediately when suddenly taken out of their airless medium, and that the result might be different if they had been gradually brought under the action of air in small quantities at a time.
We are compelled here to admit that vibrios frequently abound in liquids exposed to the air, and that they appropriate the atmospheric oxygen, and could not withstand a sudden removal from its influence. Must we, then, believe that such vibrios are absolutely different from those of butyric fermentations? It would, perhaps, be more natural to admit that in the one case there is an adaptation to life with air, and in the other case an adaptation to life without air; each of the varieties perishing when suddenly transferred from its habitual condition to that of the other, whilst by a series of progressive changes one might be modified into the other.〖These doubts might be easily removed by putting the matter to the test of direct experiment.〗 We know that in the case of alcoholic ferments, although these can actually live without air, propagation is wonderfully assisted by the presence of minute quantities of air; and certain experiments which we have not yet published lead us to believe that, after having lived without air, they cannot be suddenly exposed with impunity to the influence of large quantities of oxygen.
We must not forget, however, that aërobian torulae and anaërobian ferments present an example of organisms apparently identical, in which, however, we have not yet been able to discover any ties of a common origin. Hence we are forced to regard them as a distinct species; and so it is possible that there may likewise be aërobian and anaërobian vibrios without any transformation of the one into the other.
The question has been raised whether vibrios, especially those which we have shown to be the ferment of butyric and many other fermentations, are in their nature, animal or vegetable. M. Ch. Robin attaches great importance to the solution of this question, of which he speaks as follows:〖ROBIN, Sur la nature des fermentations, &c. (Journal de l'Académie et de la Physiologie, July and August, 1875, p. 386).〗 “The determination of the nature, whether animal or vegetable, of organisms, either as a whole or in respect to their anatomical parts, assimilative or reproductive, is a problem which has been capable of solution for a quarter of a century. The method has been brought to a state of remarkable precision, experimentally, as well as in its theoretical aspects, since those who devote their attention to the organic sciences consider it indispensable in every observation and experiment to determine accurately, before anything else, whether the object of their study is animal or vegetable in its nature, whether adult or otherwise. To neglect this is as serious an omission for such students as for chemists would be the neglecting to determine whether it is nitrogen or hydrogen, urea or stearine, that has been extracted from a tissue, or which it is whose combinations they are studying in this or that chemical operation. Now, scarcely any one of those who study fermentations, properly so-called, and putrefactions, ever pay any attention to the preceding data.... Among the observers to whom I allude, even M. Pasteur is to be found, who, even in his most recent communications, omits to state definitely what is the nature of many of the ferments which he has studied, with the exception, however, of those which belong to the cryptogamic group called torulaceae. Various passages in his work seem to show that he considers the cryptogamic organisms called bacteria, as well as those known as vibrios, as belonging to the animal kingdom (see Bulletin de l'Académie de Médecine, Paris, 1875, pp. 249, 251, especially 256, 266, 267, 289, and 290). These would be very different, at least physiologically, the former being anaërobian, that is to say, requiring no air to enable them to live, and being killed by oxygen, should it be dissolved in the liquid to any considerable extent.”
We are unable to see the matter in the same light as our learned colleague does; to our thinking, we should be labouring under a great delusion were we to suppose “that it is quite as serious an omission not to determine the animal or vegetable nature of a ferment as it would be to confound nitrogen with hydrogen or urea with stearine.” The importance of the solutions of disputed questions often depends on the point of view from which these are regarded. As far as the result of our labours is concerned, we devoted our attention to these two questions exclusively: 1. Is the ferment, in every fermentation properly so called, an organized being? 2. Can this organized being live without air? Now, what bearing can the question of the animal or vegetable nature of the ferment, of the organized being, have upon the investigation of these two problems? In studying butyric fermentation, for example, we endeavoured to establish these two fundamental points; 1. The butyric ferment is a vibrio. 2. This vibrio may dispense with air in its life, and, as a matter of fact, does dispense with it in the act of producing butyric fermentation. We did not consider it at all necessary to pronounce any opinion as to the animal or vegetable nature of this organism, and, even up to the present moment, the idea that vibrio is an animal and not a plant is in our minds, a matter of sentiment rather than of conviction.
M. Robin, however, would have no difficulty in determining the limits of the two kingdoms. According to him, “every variety of cellulose is, we may say, insoluble in ammonia, as also are the reproductive elements of plants, whether male or female. Whatever phase of evolution the elements which reproduce a new individual may have reached, treatment with this reagent, either cold or raised to boiling, leaves them absolutely intact under the eyes of the observer, except that their contents, from being partially dissolved, become more transparent. Every vegetable whether microscopic or not, every mycelium and every spore, thus preserves in its entirety its special characteristics of form, volume and structural arrangements; whilst in the case of microscopic animals, or the ova and microscopic embryos of different members of the animal kingdom, the very opposite is the case.”
We should be glad to learn that the employment of a drop of ammonia would enable us to pronounce an opinion with this degree of confidence on the nature of the lowest microscopic beings; but is M. Robin absolutely correct in his assumptions? That gentleman himself remarks that spermatozoa, which belong to animal organisms, are insoluble in ammonia, the effect of which is merely to make them paler. If a difference of action in certain reagents, in ammonia, for example, were sufficient to determine the limits of the animal and vegetable kingdoms, might we not argue that there must be a very great and natural difference between moulds and bacteria, inasmuch as the presence of a small quantity of acid in the nutritive medium facilitates the growth and propagation of the former, whilst it is able to prevent the life of bacteria and vibrios? Although as is well known, movement is not an exclusive characteristic of animals, yet we have always been inclined to regard vibrios as animals, on account of the peculiar character of their movements. How greatly they differ in this respect from the diatomacae, for example! When the vibrio encounters an obstacle it turns, or after assuring itself by some visual effort or other that it cannot overcome it, it retraces its steps. The colpoda—undoubted infusoria—behave in an exactly similar manner. It is true one may argue that the zoospores of certain cryptogamia exhibit similar movements; but do not these zoospores possess as much of an animal nature as do the spermatozoa? As far as bacteria are concerned, when, as already remarked, we see them crowd round a bubble of air in a liquid to prolong their life, oxygen having failed them everywhere else, how can we avoid believing that they are animated by an instinct for life, of the same kind that we find in animals? M. Robin seems to us to be wrong in supposing that it is possible to draw any absolute line of separation between the animal and vegetable kingdoms. The settlement of this line however, we repeat again, no matter what it may be, has no serious bearing upon the questions that have been the subject of our researches.
In like manner the difficulty which M. Robin has raised in objecting to the employment of the word germ, when we cannot specify whether the nature of that germ is animal or vegetable, is in many respects an unnecessary one. In all the questions which we have discussed, whether we were speaking of fermentation or spontaneous generation, the word germ has been used in the sense of origin of living organism. If Liebig, for example, said of an albuminous substance that it gave birth to ferment, could we contradict him more plainly than by replying “No; ferment is an organized being, the germ of which is always present, and the albuminous substance merely serves by its occurrence to nourish the germ and its successive generations”?
In our Memoir of 1862, on so called spontaneous generations, would it not have been an entire mistake to have attempted to assign specific names to the microscopic organisms which we met with in the course of our observations? Not only would we have met with extreme difficulty in the attempt, arising from the state of extreme confusion which even in the present day exists in the classification and nomenclature of these microscopic organisms, but we should have been forced to sacrifice clearness in our work besides; at all events, we should have wandered from our principal object, which was the determination of the presence or absence of life in general, and had nothing to do with the manifestation of a particular kind of life in this or that species, animal or vegetable. Thus we have systematically employed the vaguest nomenclature, such as mucors, torulae, bacteria, and vibrios. There was nothing arbitrary in our doing this, whereas there is much that is arbitrary in adopting a definite system of nomenclature, and applying it to organisms but imperfectly known, the differences or resemblances between which are only recognizable through certain characteristics, the true signification of which is obscure. Take, for example, the extensive vibrio in the works of Cohn, H. Hoffmann, Hallier, and Billroth. The confusion which prevails here is very great, although we do not of course by any means place these different works on the same footing as regards their respective merits.
M. Robin is, however, right in recognizing the impossibility of maintaining in the present day, as he formerly did, “That fermentation is an exterior phenomenon, going on outside cryptogamic cells, a phenomenon of contact. It is probably,” he adds, “an interior and molecular action at work in the innermost recesses of the substance of each cell.” From the day when we first proved that it is possible for all organized ferments, properly so called, to spring up and multiply from their respective germs, sown, whether consciously or by accident, in a mineral medium free from organic and nitrogenous matters other than ammonia, in which medium the fermentable matter alone is adapted to provide the ferment with whatever carbon enters into its composition, from that time forward the theories of Liebig, as well as Berzelius, which M. Robin formerly defended, have had to give place to others more in harmony with facts. We trust that the day will come when M. Robin will likewise acknowledge that he has been in error on the subject of the doctrine of spontaneous generation, which he continues to affirm, without adducing any direct proofs in support of it, at the end of the article to which we have been here replying.
We have devoted the greater part of this chapter to the establishing with all possible exactness the extremely important physiological fact of life without air, and its correlation to the phenomena of fermentations properly so called—that is to say, of those which are due to the presence of microscopic cellular organisms. This is the chief basis of the new theory that we propose for the explanation of these phenomena. The details into which we have entered were indispensable on account of the novelty of the subject no less than on account of the necessity we were under of combating the criticisms of the two German naturalists, Drs. Oscar Brefeld and Traube, whose works had cast some doubts on the correctness of the facts upon which we had based the preceding propositions. We have much pleasure in adding that at the very moment we were revising the proofs of this chapter, we received from M. Brefeld an essay, dated Berlin, January, 1876, in which, after describing his later experimental researches, he owns with praiseworthy frankness that Dr. Traube and he were both of them mistaken. Life without air is now a proposition which he accepts as perfectly demonstrated. He has witnessed it in the case of mucor racemosus and has also verified it in the case of yeast. “If,” he says, “after the results of my previous researches, which I conducted with all possible exactness, I was inclined to consider Pasteur's assertion as inaccurate and to attack them, I have no hesitation now in recognizing them as true, and in proclaiming the service which Pasteur has rendered to science in being the first to indicate the exact relation of things in the phenomenon of fermentation.” In his later researches, Dr. Brefeld has adopted the method which we have long employed for demonstrating the life and multiplication of butyric vibrios in the entire absence of air, as well as the method of conducting growths in mineral media associated with fermentable substance. We need not pause to consider certain other secondary criticisms of Dr. Brefeld. A perusal of the present work will, we trust, convince him that they are based on no surer foundation than were his former criticisms.
To bring one's self to believe in a truth that has just dawned upon one is the first step towards progress; to persuade others is the second. There is a third step, less useful perhaps, but highly gratifying nevertheless, which is, to convince one's opponents.
We therefore, have experienced great satisfaction in learning that we have won over to our ideas an observer of singular ability, on a subject which is of the utmost importance to the physiology of cells.